ERGEBNISSE UND GRUNDLAGEN VON IMMUNREAKTIONEN BEI
ARTHOPDEN
Die Grundlage eines jeden Immunsystems ist die Fähigkeit zur
Unterscheidung von Selbst und Fremd.
Nun ist es wichtig zu verstehen, daß bei Evertebraten Antikörper
und MHC (major histocompatibility complex) nicht gefunden wurden, also andere
Mechanismen der Fremderkennung zugrunde liegen müssen.
In Frage kommen hier physikalische Eigenschaften einer Oberfläche
wie deren Ladung oder Hydrophobizität, physikalisch-)chemische
Eigenschaften bestimmter mikrobieller Stoffen wie z.B. LPS oder
ß-1,3-Glucanen.
Die Hauptakteure bei den Immunreaktionen der Arthropoden sind
die Haemocyten. Auch heute noch werden sie vorwiegend aufgrund ihrer
Form und Einschlüsse und ihres in vitro-Verhaltens unterschieden
und angesprochen.
Die prominentesten Typen sind Prohaemocyten, Plasmatocyten, Granuläre
Haemocyten und Sphaerulocyten. Allerdings ist die Nomenklatur
nicht für alle Arthropodengruppen einheitlich, und selbst innerhalb
der Insekten (Tracheaten) fällt eine Homologisierung der Haemocyten-Typen
schwer.
Aber es werden zunehmen mehr monoklonale Antikörper gefunden, die bestimmte
Haemocytentypen auch in verschiedenen Taxa markieren können.
Wenn nun Haemocyten durch den Kontakt mit einer Oberfläche stimuliert
worden sind, kommt es zu u.U. rasanten Veränderungen der im nativen
Zustand meist sphaerisch, ovoiden oder discoiden Zellen.
Die Augenfälligsten Aktivitäten sind Degranulation,
Anheftung und Spreiten, Phagocytose, Aggregation
und Noduli bzw. Kapselbildung.
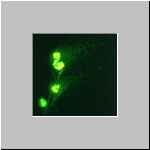
Eine effektvolle Darstellung der Degranulation von in Kulturmedium
auf einem Objektträger gespreiteten Lithobius-Haemocyten ist
hier mit fluoreszenzfarbstoff-gekoppeltem WGA (Wheat Germ Agglutinin)
gelungen.
Die Zellen wurde vor der Färbung mit Methanol fixiert.
Durch das Farbstoff-gekoppelte Lektin WGA wurde genau das exocytierte, von
den Zellen wegdriftende Material und das noch in den Grana befindliche
Ausgangsmaterial, deutlich markiert.
Das exocytierte Material unterliegt im nicht fixiertem Zustand offenbar
einer Polymerisation, ähnlich der des Fibrins.
Die Degranulation der Haemocyten dient also zum einen dazu, eine Koagulation
und damit einen Wundverschluß herbeizuführen.
Allerdings können durch Degranulation auch antibakterielle Substanzen
freigesetzt werden, ich gehe später noch einmal darauf ein.
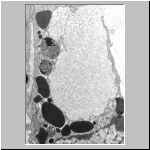
Das TEM-Bild einer Scolopendra-Haemocyte verdeutlicht hier nocheinmal
wie sich der Inhalt spezieller Grana verändert bevor es als faserig
strukturiertes Material exocytiert wird.
Kommen wir nun zum Anhaften und Spreiten der Haemocyte.
Dies ist auf einer inerten Oberfläche von deren physikalischen Eigenschaften
abhängig. Allerdings sind die allermeisten artifiziellen den eigenen
Oberflächen, etwa den Basallaminae, relativ unähnlich, werden also
von den Haemocyten fast immer als fremd erkannt.
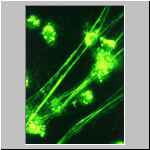
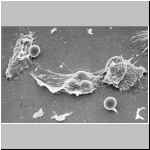
Auf einem Glasobjektträgern mit Kulturmedium haben sich diese
Lithobius-Haemocyten daher schnell gespreiten; die Färbung –
ein Nachweis von PO-Aktivität – ist hier nicht wichtig. Gut zu
erkennen ist, daß die verschiedenen Haemocytentypen unterschiedlich
spreitaktiv sind. Plasmatocyten spreiten in alle Richtungen, Granuläre
Haemocyten spreiten unsymmetrisch und Sphaerulocyten spreiten gar nicht oder
nur sehr wenig.
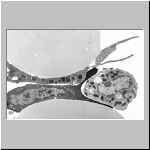
Dieses TEM-Bild zeigt an großen Latex-Kugeln gespreitete
L. forficatus-Haemocyten, die Zellen sich sehr eng an die Kugeln
angeheftet.
Und diese migrierende Plasmatocyte, die offenbar kleine Latex-Partikel
phagocytiert hat, macht deutlich, daß Spreiten in gewisser Weise
ein Versuch ist, große Objekte zu Phagocytieren; womit wir zum
nächsten Punkt kommen.